Биология в лицее
Сайт учителей биологии МБОУ Лицей № 2 г. Воронежа, РФ ![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
Site biology teachers lyceum № 2 Voronezh city, Russian Federation
|
Хромосомная теория наследственности. Сцепленное наследование генов
Мендель проследил наследование только семи пар признаков у душистого горошка. В дальнейшем многие исследователи, изучая наследование разных признаков у самых разных видов организмов, подтвердили законы Менделя. Было признано, что эти законы носят всеобщий характер. Однако позже было замечено, что у душистого горошка два признака — форма пыльцы и окраска цветков — не дают независимого распределения в потомстве и потомки остаются похожими на родителей. Постепенно таких исключений из третьего закона Менделя накапливалось всё больше. Стало ясно, что принцип независимого распределения в потомстве и свободного комбинирования распространяется не на все гены. В самом деле, у любого организма признаков очень много, а число хромосом невелико. Следовательно, в каждой хромосоме должно находиться много генов. К примеру, у человека на одну хромосому приходится в среднем более тысячи генов. Каковы же закономерности наследования генов, локализованных в одной хромосоме? Этот вопрос был изучен выдающимся американским генетиком Томасом Морганом.
Явление совместного наследования генов, локализованных в одной хромосоме, называется сцепленным наследованием, а локализация генов в одной хромосоме — сцеплением генов. Сцепленное наследование генов, локализованных в одной хромосоме, называют законом Моргана.
Все гены, входящие в одну хромосому, как вы помните, передаются по наследству совместно и составляют группу сцепления. Поскольку в гомологичных хромосомах находятся одинаковые гены, группу сцепления образуют две гомологичные хромосомы. Число групп сцепления соответствует числу хромосом в гаплоидном наборе. Так, у человека 46 хромосом — 23 группы сцепления. У дрозофилы 8 хромосом — 4 группы сцепления, у гороха 14 хромосом — 7 групп сцепления.
Предположим, что два гена — A и B находятся в одной хромосоме и организм, взятый для скрещивания, гетерозиготен по этим генам. В анафазе первого мейотического деления гомологичные хромосомы расходятся в разные клетки, и образуются два сорта гамет вместо четырёх, как должно было бы быть при дигибридном скрещивании в соответствии с третьим законом Менделя. При скрещивании с гомозиготным организмом, рецессивным по обоим генам — aa и bb (анализирующее скрещивание), получается расщепление 1 : 1 вместо ожидаемого при дигибридном анализирующем скрещивании 1 : 1 : 1 : 1.
Такое отклонение от независимого распределения означает, что гены, локализованные в одной хромосоме, наследуются совместно. Если при анализирующем скрещивании в случае сцепленного наследования в потомстве дигетерозиготного организма наблюдается расщепление в соотношении 1 : 1, то говорят о полном сцеплении генов. Это означает, что гены A и B (а также a и b) расположены в хромосоме непосредственно друг за другом и кроссинговер между ними практически невозможен. Обмен другими участками в данной паре гомологичных хромосом никак не скажется на наследовании генов A и B (a и b).
Сцепление генов. Наследование генов, расположенных в хромосоме друг за другом. (На схеме скрещивания для удобства гены расположены иначе, чем на рисунке слева, где показано физическое расположение генов в хромосоме. Конечно, для наглядности размеры генов сильно преувеличены)
Однако при анализе наследования сцепленных генов было обнаружено, что в некотором проценте случаев, строго определённом для каждой пары генов, сцепление может нарушаться. Если в результате кроссинговера в некоторых клетках происходит обмен участками хромосом между генами A и B, появляются гаметы Ab и aB, и в потомстве образуются четыре группы фенотипов, как при свободном комбинировании генов. Различие заключается в том, что числовое отношение фенотипов в подавляющем большинстве случаев не соответствует отношению 1 : 1 : 1 : 1, установленному для дигибридного анализирующего скрещивания.
Рассматривая как раз такой редкий случай, как, например, характер наследования генов А и C (a и c), находящихся на разных концах плеч хромосом, мы будем наблюдать иную картину. Поскольку вероятность кроссинговера для целой хромосомы составляет 50%, то количество гамет, при образовании которых кроссинговера не было, составит лишь 50% (по 25% AC и ac). Эти гаметы получили название некроссоверных. В остальных 50% случаев появляются половые клетки с перекомбинированными генами (по 25% aC и Ac) — кроссоверные гаметы. Анализирующее скрещивание с таким организмом даст четыре группы потомков в соотношении 1 : 1 : 1 : 1, как при независимом наследовании.
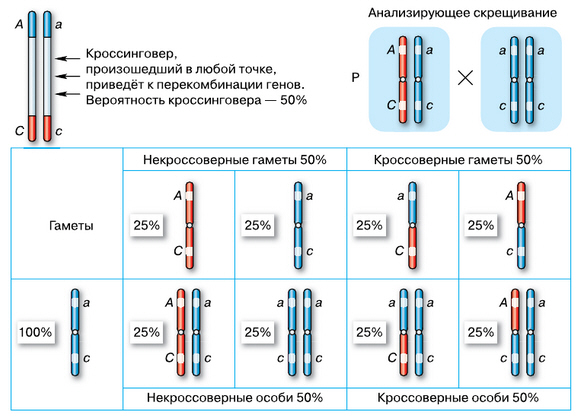 Сцепление генов. Наследование генов, расположенных на разных концах плеч хромосомы. (На схеме скрещивания для удобства гены расположены иначе, чем на рисунке слева, где показано физическое расположение генов в хромосоме. Конечно, для наглядности размеры генов сильно преувеличены)
В третьей ситуации разместим ген A (a) на конце плеча хромосомы, а ген D (d) локализуем в её геометрической середине. В этом случае вероятность кроссинговера для участка хромосомы между этими генами составит 50% : 2 = 25% . Обмены участками в области хромосомы за геном D (d), как и в первом случае (ниже гена B), не окажут влияния на наследование генов данной пары. Таким образом, в 75% случаев образования гамет перекомбинирования не произойдёт, и некроссоверные гаметы будут выглядеть следующим образом: 37,5% AD и 37,5% ad. В 25% — кроссинговер «разлучит» гены A и D (a и d) — возникнут кроссоверные гаметы: 12,5% aD и 12,5% Ad. Сходное числовое соотношение получится и при анализирующем скрещивании в потомстве.
Сцепление генов. Наследование генов, один из которых расположен на конце хромосомы, а другой — на её геометрической середине. (На схеме скрещивания для удобства гены расположены иначе, чем на рисунке слева, где показано физическое расположение генов в хромосоме. Конечно, для наглядности размеры генов сильно преувеличены)
Таким образом, сцепление генов может быть полным и неполным. Причиной нарушения сцепления служит кроссинговер — перекрёст хромосом в профазе первого мейотического деления. Чем дальше друг от друга расположены гены в хромосоме, тем выше вероятность перекрёста между ними и тем больше процент гамет с перекомбинированными генами, а следовательно, и больший процент особей, отличающихся от родителей. В генетике принято оценивать расстояние между генами, расположенными в одной хромосоме, в процентах гамет, при образовании которых в результате кроссинговера произошла перекомбинация генов в гомологичных хромосомах. За единицу расстояния между генами, находящимися в одной хромосоме, принят 1% кроссинговера. Такая единица в честь Т. Моргана получила название морганиды. Следовательно, в нашем первом примере при полном сцеплении генов расстояние между ними составляет 0 морганид, во втором — 50, а в последнем — 25 морганид. Рассмотрим конкретный пример. У дрозофилы гены, определяющие цвет тела и размеры крыльев, расположены в одной хромосоме. Доминантный ген, обусловливающий развитие серой окраски тела, — A, преобладает над геном a, определяющим тёмную окраску. Ген B приводит к появлению нормальных, а b — рудиментарных крыльев у насекомого. Если скрестить гомозиготную мушку-дрозофилу, имеющую серое тело и нормальные крылья, с насекомым, обладающим тёмной окраской тела и зачаточными крыльями, то в первом поколении гибридов все мухи будут серыми с нормальными крыльями. Это гетерозиготы по двум парам аллельных генов, причём ген A, определяющий серую окраску брюшка, расположен в одной хромосоме с геном нормальных крыльев, а рецессивные гены этих пар находятся в другой гомологичной хромосоме.
При анализирующем скрещивании в потомстве дигетерозиготы в значительно меньшем количестве, чем особей, схожих по фенотипу с родителями, появляются серые мухи с рудиментарными крыльями и тёмноокрашенные мухи с нормальными крыльями. Возникновение организмов с такими комбинациями признаков в незначительном количестве (по 8,5% каждого типа) обусловлено нарушением сцепления генов. Таким образом, в приведённом примере расстояние между генами составило 17 морганид.
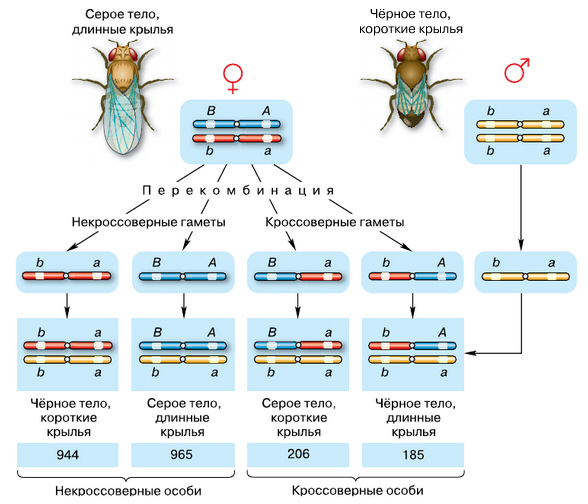 Схема опыта Т. Моргана
|
Меню сайтаСтатистикаОнлайн всего: 1 Гостей: 1 Пользователей: 0 Наш опросМини-чатПоиск
|




6+ Информация, размещенная на страницах сайта "Биология в лицее", относится к информационной продукции, допускаемой к обороту для детей, достигших возраста шести лет в соответствии со ст. 8 ФЗ №436 от 29.12.2010 г.