Биология в лицее
Сайт учителей биологии МБОУ Лицей № 2 г. Воронежа, РФ ![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
Site biology teachers lyceum № 2 Voronezh city, Russian Federation
|
Мейоз Закономерности прохождения мейоза в мужских и женских половых клетках по существу одинаковы. Сущность мейоза состоит в том, что каждая половая клетка получает одинарный — гаплоидный набор хромосом. Вместе с тем мейоз — это процесс, в результате которого создаются новые комбинации генов путём сочетания разных материнских и отцовских хромосом. Перекомбинирование наследственных задатков возникает, кроме того, и в результате обмена участками между гомологичными хромосомами, происходящего в мейозе.
Мейоз включает два последовательных, следующих друг за другом практически без перерыва деления. Как и при митозе, в каждом мейотическом делении выделяют четыре стадии: профазу, метафазу, анафазу и телофазу.
Первое мейотическое деление. Первое мейотическое деление носит название редукционного (от лат. reductio — уменьшение), так как после его завершения в дочерних клетках оказывается вдвое меньше хромосом, чем в исходной.
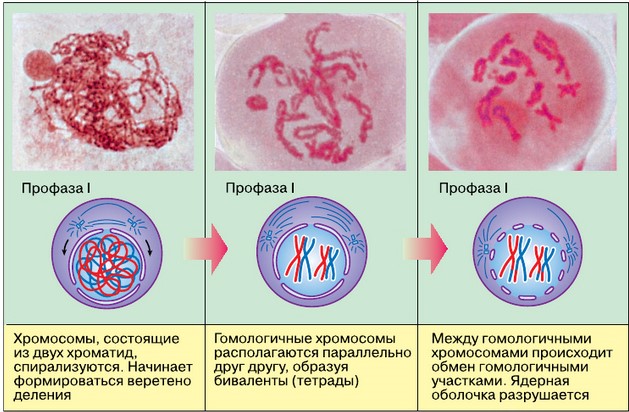
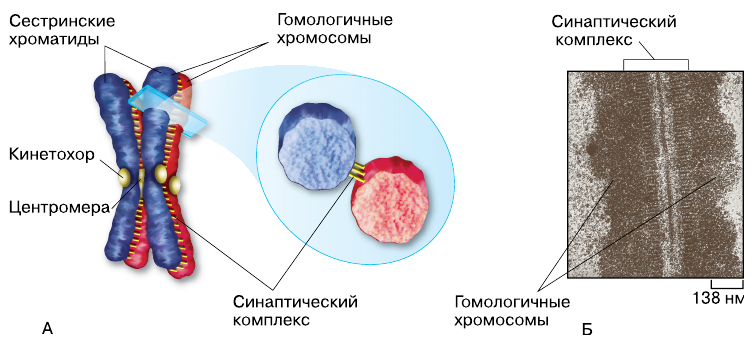
Профаза I начинается спирализацией хромосом, которые видны в световой микроскоп как тонкие длинные нити. Уже на этом этапе при большом увеличении микроскопа можно различить, что каждая хромосома состоит из двух хроматид, соединённых между собой в области центромеры. Затем гомологичные хромосомы сближаются, каждая точка одной хромосомы совмещается с соответствующей точкой другой гомологичной хромосомы. Этот процесс точного и тесного сближения гомологичных хромосом в мейозе называется конъюгацией. В процессе конъюгации гомологичные хромосомы, каждая из которых состоит из двух хроматид, сближаются на расстояние, не превышающее 120 нм. Но они не просто лежат рядом, а удерживаются одна около другой благодаря образованию хроматидами каждой гомологичной хромосомы белковых нитей с утолщениями на свободном конце (синаптический комплекс). Нити обеих хромосом соединяются друг с другом способом, напоминающим застёжку-молнию.
Конъюгация гомологичных хромосом: А — схема образования синаптического комплекса; Б — электронная микрофотография синаптического комплекса
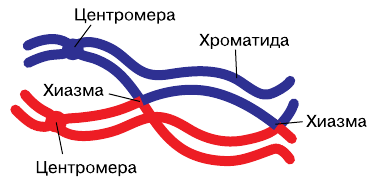 Благодаря таким мостикам гомологичные хромосомы могут долгое время находиться в сближенном (конъюгированном) состоянии. Пара конъюгированных хромосом образует бивалент (от лат. bi — двойной) или тетраду (от греч. tetrados — четвёрка). Благодаря таким мостикам гомологичные хромосомы могут долгое время находиться в сближенном (конъюгированном) состоянии. Пара конъюгированных хромосом образует бивалент (от лат. bi — двойной) или тетраду (от греч. tetrados — четвёрка).Схема кроссинговера, при котором обмениваются участками несестринские хроматиды
В дальнейшем между хромосомами, составляющими бивалент, может произойти обмен одинаковыми, или гомологичными, т. е. содержащими одни и те же гены, участками. Такой процесс носит название кроссинговера (от англ. crossing-over — перекрёст).
Вероятность кроссинговера для каждой хромосомы составляет 50%. При этом участками обмениваются две рядом лежащие несестринские хроматиды, входящие в состав разных гомологичных хромосом. Обмен участками включает два эпизода: разрыв хроматид на одном уровне и соединение центральной части плеча одной хроматиды с периферической частью плеча другой несестринской хроматиды. В местах кроссинговера формируются физические перекрёсты — хиазмы (от лат. hiazma — перекрёст).
В результате кроссинговера каждая из гомологичных хромосом оказывается состоящей из одной хроматиды с неизменённым набором генов и второй — с перекомбинированными генами. Таким образом, в составе бивалента в результате обмена участками все хроматиды оказываются разными по комбинациям вариантов аллельных генов.
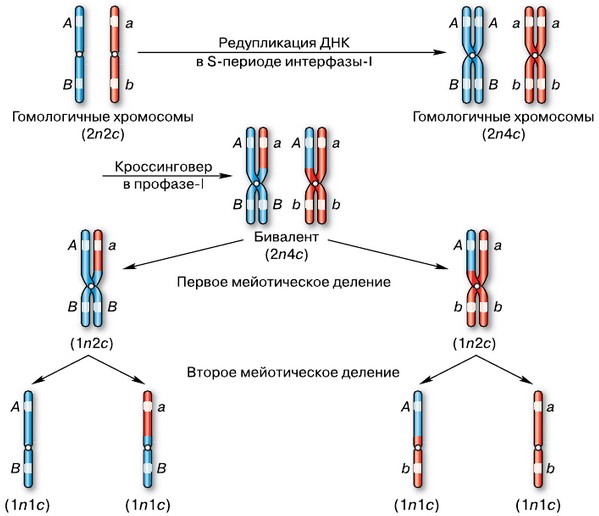 Генетическая схема мейоза. Все дочерние клетки различаются по комбинации генов
К концу профазы между гомологичными хромосомами возникают силы отталкивания. Вначале они проявляются в области центромер, а затем в других участках. Хромосомы остаются связанными между собой только в местах кроссинговера — хиазмах. По мере усиления спирализации возрастают и силы отталкивания. Хиазмы смещаются к концам плеч хромосом, где образуются терминальные (конечные) хиазмы.
При отсутствии кроссинговера между гомологичными хромосомами силы отталкивания как бы «расстёгивают» застёжку-молнию в направлении от центромеры к концам плеч хромосом.
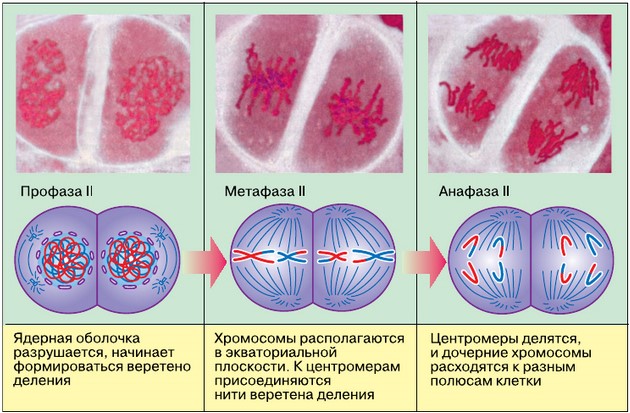
В метафазе I спирализация хромосом достигает максимума. Конъюгированные хромосомы располагаются по экватору клетки, причём в плоскости экватора лежат теломерные участки хромосом, соединённые терминальными хиазмами, а центромеры гомологичных хромосом обращены к разным полюсам клетки. К ним прикрепляются микротрубочки веретена деления.
В анафазе I плечи гомологичных хромосом окончательно разделяются в результате разрыва терминальных хиазм, и гомологичные хромосомы из бивалента расходятся к различным полюсам клетки. Следовательно, из каждой пары гомологичных хромосом в дочернюю клетку попадает только одна хромосома. Число хромосом уменьшается в два раза, и хромосомный набор становится гаплоидным. Однако каждая хромосома состоит из двух хроматид, т. е. по-прежнему содержит удвоенное количество ДНК, и, следовательно, хромосомный набор клетки после завершения первого мейотического деления будет 1n2c.
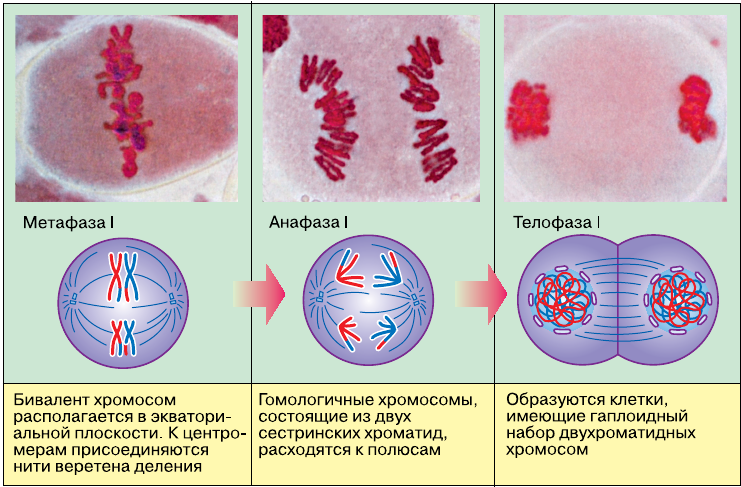 В телофазе I на короткое время образуется ядерная оболочка.
Интерфаза II. Поскольку отдельные хромосомы гаплоидных дочерних клеток продолжают оставаться удвоенными, во время интерфазы между первым и вторым делениями мейоза редупликации ДНК не происходит. Идёт репаративный синтез ДНК, направленный на ликвидацию возможных нарушений структуры ДНК, возникших в результате кроссинговера.
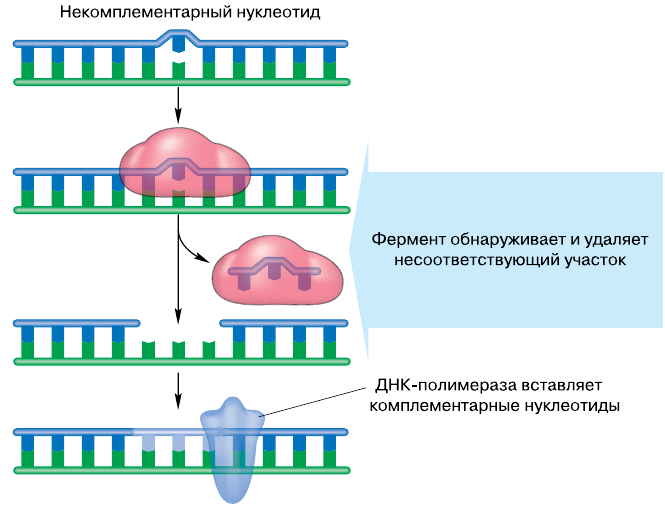 Репарация ДНК Репарация ДНККлетки, образовавшиеся в результате первого деления созревания, отличаются по составу отцовских и материнских хромосом и, следовательно, по набору генов. Например, все клетки человека, в том числе первичные половые клетки, содержат 46 хромосом. Из них 23 получены от отца и 23 от матери. После первого мейотического деления в сперматоциты и овоциты попадает только по 23 хромосомы — по одной хромосоме из каждой пары гомологичных хромосом. Однако вследствие случайности расхождения отцовских и материнских хромосом в анафазе I образующиеся клетки получают самые разнообразные комбинации родительских хромосом. Например, в одной из них может оказаться 3 отцовских и 20 материнских хромосом, в другой — 10 отцовских и 13 материнских, в третьей — 20 отцовских и 3 материнских и т. д. Число возможных комбинаций очень велико и составляет 223 варианта. Если учесть ещё обмен гомологичными участками хромосом в профазе I деления мейоза, то вполне очевидно, что каждая образующаяся клетка генетически уникальна, так как несёт свой неповторимый набор генов. Следовательно, мейоз — основа комбинативной генотипической изменчивости.
Второе мейотическое деление называют эквационным (от лат. aequatio — уравнивание). Оно в общем протекает так же, как обычное митотическое деление, с той лишь разницей, что делящаяся клетка гаплоидна (1n2c).
Таким образом, сущность периода созревания состоит в том, что в половых клетках путём двукратного мейотического деления количество хромосом уменьшается вдвое, а количество ДНК — вчетверо. Биологический смысл второго мейотического деления заключается в приведении количества ДНК в соответствие хромосомному набору.
< Предыдущая страница "Развитие половых клеток (гаметогенез)" |
Меню сайтаСтатистикаОнлайн всего: 1 Гостей: 1 Пользователей: 0 Наш опросМини-чатПоиск
|
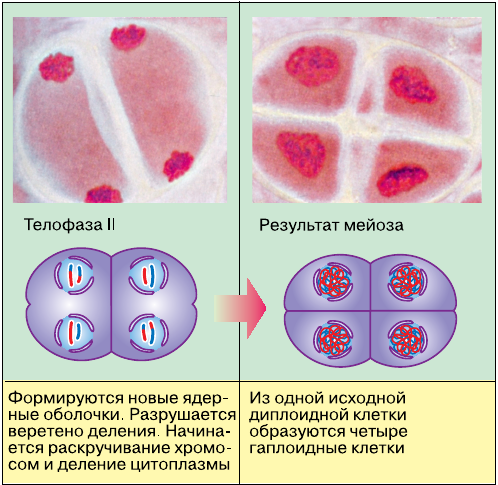 В анафазе II центромеры, соединяющие сестринские хроматиды в каждой хромосоме, делятся. Хроматиды, как и в митозе, с этого момента становятся самостоятельными дочерними хромосомами и начинают движение к разным полюсам клетки. С завершением телофазы II заканчивается и весь процесс мейоза. В результате из исходной первичной половой клетки образовались четыре гаплоидные клетки с хромосомным набором 1n1c.
В анафазе II центромеры, соединяющие сестринские хроматиды в каждой хромосоме, делятся. Хроматиды, как и в митозе, с этого момента становятся самостоятельными дочерними хромосомами и начинают движение к разным полюсам клетки. С завершением телофазы II заканчивается и весь процесс мейоза. В результате из исходной первичной половой клетки образовались четыре гаплоидные клетки с хромосомным набором 1n1c.


6+ Информация, размещенная на страницах сайта "Биология в лицее", относится к информационной продукции, допускаемой к обороту для детей, достигших возраста шести лет в соответствии со ст. 8 ФЗ №436 от 29.12.2010 г.